
Dr. Gerardo Barroso Director Científico
Dra. Alinne Colín Médico Asociado
Centro de reproducción nascere
 INTRODUCCIÓN
INTRODUCCIÓN
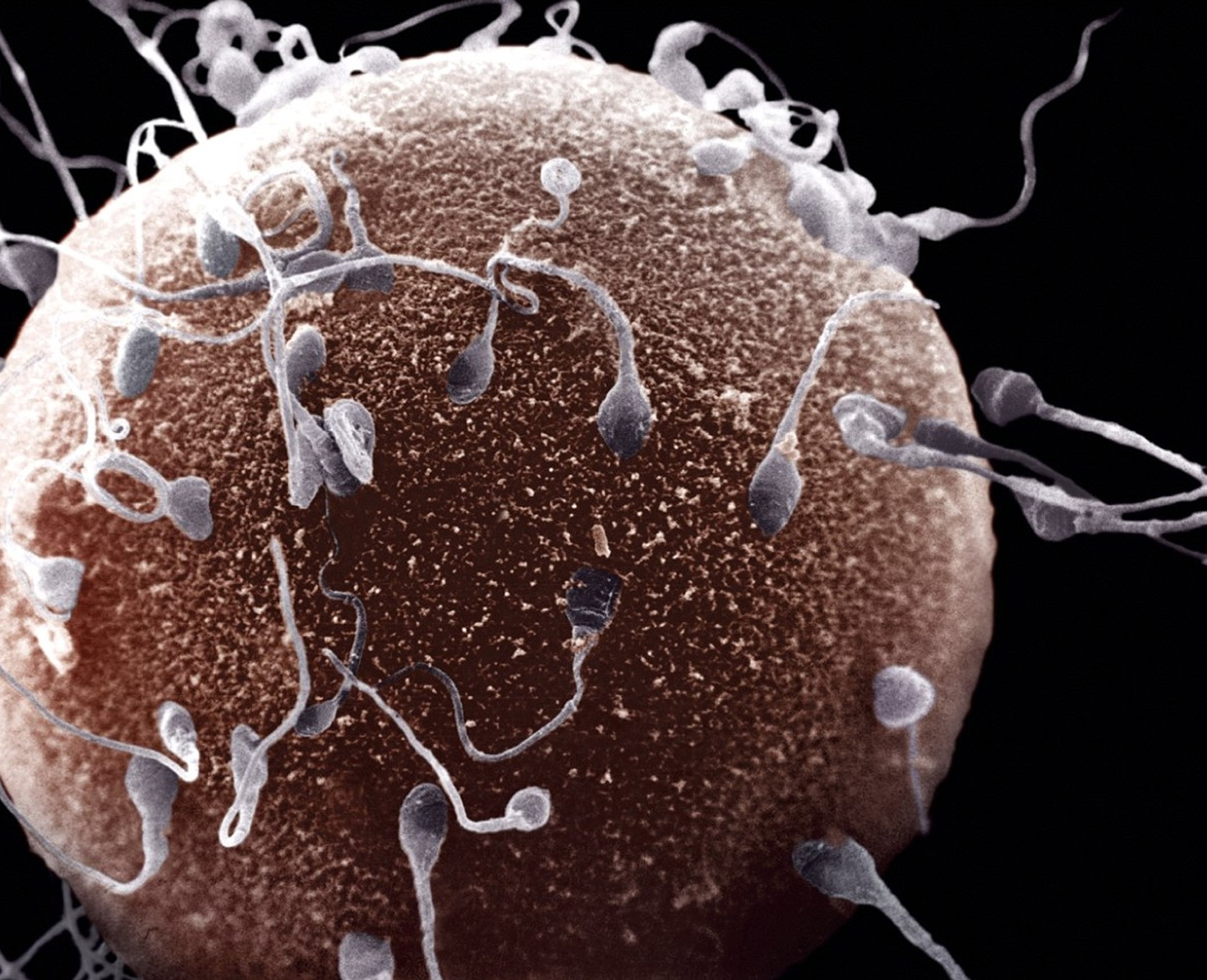
Al hablar del factor masculino, inmediatamente nos viene a la mente la palabra “espermatozoide”, y si nos preguntaran acerca del concepto de éste, probablemente responderíamos que es el gameto masculino que aporta la información genética paterna para la formación de un nuevo individuo, lo cual puede resultar sencillo de leer; sin embargo, los procesos que deben sincronizarse para llegar a la fertilización resultan tremendamente complejos, por lo que en este breve capítulo describiremos los procesos más relevantes y algunas de las alternativas terapéuticas en pro de soslayar las patologías del espermatozoide.
Para dimensionar la importancia de esta pequeña célula, se debe mencionar que, actualmente, más de ochenta millones de personas en el mundo padecen de infertilidad, y contrariamente a lo que tradicionalmente se pensaría, cerca del 50 por ciento de los casos son resultado de alteraciones en el factor masculino, por lo que entender la función del gameto masculino nos ha ayudado en la mejora de los procesos de reproducción asistida.
En esa forma, las dos últimas décadas han sido decisivas en el devenir tecnológico en materia de alta tecnología reproductiva; los procesos de alta complejidad, como es la fertilización in-vitro (FIV) no sólo han venido a dar solución terapéutica a los problemas de infertilidad, sino también han servido como herramientas diagnósticas en los mecanismos inherentes a la fertilización humana y su disfunción, lo cual ha brindado información suficiente para enmarcar al gameto masculino como desencadenante de eventos biomoleculares y de señalización que impactarán en el desarrollo humano. Más allá de esto, hoy tenemos la capacidad de evaluar cada una de las etapas del desarrollo humano, desde la interacción óvulo-espermatozoide.
LA ZAGA DEL ESPERMATOZOIDE
Pero, ¿cuál es largo camino por el que esta pequeña célula debe pasar para lograr crear una nueva vida?
Las respuestas a esta interrogante inician al dar una mirada atrás y remontarnos al mismo desarrollo embrionario, ya que desde la fusión del óvulo (gameto femenino) y el espermatozoide, se define si el material genético constituirá un producto femenino con cariotipo XX o un producto masculino con cariotipo XY (ambos normales); en caso de ser así, el gen SRY será el responsable de esta diferenciación, y así devendrá la formación de tejido testicular y, con éste, el desarrollo espermático.
Esto suena sencillo; sin embargo, para que este proceso pueda llevarse a cabo, es necesario un engranaje perfecto en el desarrollo anatómico y endócrino, y es así como un producto XY llegará al mundo, para lo cual será necesario que todos los mecanismos se mantengan trabajando de forma perfectamente sincronizada, hasta la pubertad, cuando el actuar de las hormonas hará que se inicie la madurez de estas células germinales sexuales (espermatogénesis), las cuales serán renovadas aproximadamente cada tres meses.
PRINCIPALES ALTERACIONES EN EL GAMETO MASCULINO
Del breve párrafo anterior, se derivan una serie de patologías que pueden tener como fatal desenlace la falta de formación y/o función del espermatozoide. Veamos cómo:
i) En caso de existir algún defecto durante el engranaje cromosómico, éste puede resultar en un embrión sin número cromosómico normal (ya sea 46XX o 46 XY), lo cual nos dará alteraciones tan aparentes como son los defectos anatómicos en los genitales externos; sin embargo, existen…
ii) Defectos genéticos (como son microdeleciones del cromosoma Y, que ocurren más frecuentemente en el brazo largo del cromosoma Y, en un área particular en la región del factor de azoospermia o AZF, resultando en fallas variadas de la espermatogénesis. Las alteraciones más frecuentes son deleciones en las aéreas AZFa y AZFb. Los dos genes más importantes presentes en la región AZFa son USP9Y y DBY, y sus deleciones desencadenan el síndrome de células de Sertoli aisladas. Deleciones en la región AZFb determinan arresto de la espermatogénesis en el estadio de espematocitos, y el gen más importante en este nivel es RBMY. Por su parte, las microdeleciones parciales/totales de la región AZFc resultan en una gama variada de fenotipos, desde oligozoospermia a azoospermia), que resultan físicamente imperceptibles, y se vuelven evidentes cuando el paciente acude por deseo reproductivo.
iii) Los defectos en la vía genital masculina no sólo pueden ser resultado de alteraciones de los cromosomas sexuales, ya que existen patologías, como la fibrosis quística, que pueden dar como resultado alteraciones (ausencia de túbulos seminíferos) que darán como resultado infertilidad.
iv) Sobrepasando estas alteraciones, el espermatozoide enfrenta en su camino obstáculos propios del desarrollo del individuo, como la presencia de infecciones de la infancia, que pueden poner en peligro el desarrollo de esta célula ( como es la parotiditis).
v) Sin dejar de lado las causas traumáticas (golpes) que pueden afectar este proceso.
vi) Logrando sobrepasar todo lo anterior, aún nos queda el poder lograr un estímulo hormonal perfecto, tanto de hormonas hipotalámicas (hormona liberadora de gonadotropinas GnRH), como de hipofisiarias (hormona folículo estimulante FSH y luteinizante LH), lo cual permitirá un nivel adecuado de testosterona y, con todo lo anterior, un desarrollo espermático adecuado.
vii) Enlistando lo anterior, pensaríamos que ya son suficientes obstáculos en la zaga del espermatozoide, pero aún quedan otros, como son las patologías oncológica con afección testicular, y…
viii) El propio estilo de vida (someterse a ambientes con temperaturas extremas (sauna, vapor, actividad laboral que así lo requiera); el tabaquismo, obesidad o la misma edad), que son factores descritos como responsables de alteración en la calidad espermática.
Revisando lo anterior, la siguiente interrogante es: “¿Cómo damos solución a estas alteraciones, para lograr la fertilización y el desarrollo de un embrión sano?”
ALTERNATIVAS DIAGNÓSTICAS Y TERAPÉUTICAS
Para ello, es necesario remontarnos a las bases del análisis seminal (espermatobioscopia), que constituye la herramienta de primera instancia en la evaluación del factor masculino, en donde los parámetros de mayor relevancia son:
i) La cuenta espermática (se considera normal si ésta es igual a / o sobrepasa los 20 millones de células). Las alteraciones de este parámetro se conocen como oligozospermia, y su extremo, la azoospermia.
ii) La motilidad (idealmente con más del 50 por ciento de fracción con adecuada motilidad). Sus alteraciones: espectro de leve, moderada o severa de la astenozospermia.
iii) Por último, tenemos la morfología (parámetro con mayor discordancia de acuerdo al criterio utilizado. La Organización Mundial de la Salud considera como normal 4 por ciento; los criterios estrictos de Kruger, 14 por ciento). Nos referimos a alteraciones de la misma, como teratozospermia, la cual puede ir de leve a severa. Algunas décadas atrás, con este resultado sería suficiente para ofrecer un pronóstico reproductivo a la pareja infértil, y más aún con el advenimiento (hace más de 20 años) de la inyección intracitoplasmática del espermatozoide (ICSI), la cual ha venido a dar solución explícita a los problemas; antes, del espermatozoide, y ahora, de la disfunción ovocitaria y falla repetida de la fertilización.
Sin embargo, las implicaciones en el devenir reproductivo en el uso de estas técnicas en la búsqueda del embarazo han sembrado por mucho tiempo la duda en la transmisión de un gameto sano y las implicaciones que esto conlleva. Hoy, más que nunca, la alta biotecnología aplicada a los procesos moleculares en la participación e interacción del espermatozoide y óvulo preimplantatorios, nos permiten visualizar terapia que permita la selección del gameto optimizado y a su vez las implicaciones génicas y epigenéticas en el cómo y cuándo en el uso de la tecnología reproductiva.
De este modo, el análisis de la literatura revela que se han implementado varios procedimientos para obtener una selección “ideal” de espermatozoides normales y maduros para micro-inyección. Estos incluyen: gradientes de densidad, basados en la relación espermática masa: volumen, swim-up, basado en motilidad; filtración por lana de vidrio; el método zeta, basado en la carga eléctrica superficial (de poca utilización); el ácido hialurónico, basado en la presencia hipotética de un receptor específico, y, finalmente, uso de citometría de flujo, con partículas magnéticas que separan espermatozoides apoptóticos mediante la unión de Anexina V a la fosfatidilserina externalizada.
Los resultados con estas técnicas en general demuestran un incremento significativo de separación de espermatozoides en términos de morfología, motilidad, cromatina (ausencia de histonas excesivas) así como de fragmentación de ADN. Sin embargo, dados los resultados recientes que señalan la presencia de ADN fragmentado aun en células con morfología estricta normal en pacientes con OAT, es absolutamente necesario continuar la búsqueda de métodos que pudieran “asegurar” la obtención de espermatozoides con ADN íntegro para usar en la reproducción asistida, que permitan mejorar los resultados tanto de la fertilización in vitro como de la ICSI.
REFERENCIAS:
1. Oehninger S. Place of intracytoplasmic sperm injection in management of male infertility. Lancet. 2001 Jun 30;357(9274):2068-9.
2. O’Flynn O’Brien KL, Varghese AC, Agarwal A. The genetic causes of male factor infertility: a review. FertilSteril. 2010 Jan;93(1):1-12.
3. Arslan M, Oehninger S, Kruger, TF. Clinical management of male infertility. En: Oehninger SC and Kruger TF, editors. Male Infertility: Diagnosis and Treatment, United Kingdom: Informa Health Care; 2007.p.305-318
4. Barroso G, Valdespin C, Vega E, Kershenovich R, Avila R, Avendaño C, Oehninger S. Developmental sperm contributions: fertilization and beyond. FertilSteril. 2009 Sep;92(3):835-48.
5. Aitken RJ, De Iuliis GN. Origins and consequences of DNA damage in male germ cells. Reprod Biomed Online. 2007 Jun;14(6):727-33.
6. Sa R, Souza M, Cremades N, Alves C, Silva J, Barros A. En: Oehninger SC and Kruger TF, editors. Male Infertility: Diagnosis and Treatment, United Kingdom: Informa Health Care; 2007. p.425-453
7. Stukenborg JB, Schlatt S, Simoni M, Yeung CH, Elhija MA, Luetjens CM, et al. New horizons for in vitro spermatogenesis? An update on novel three-dimensional culture systems as tools for meiotic and post-meiotic differentiation of testicular germ cells. Mol Hum Reprod. 2009 Sep;15(9):521-9.
8. Haywood M, Spaliviero J, Jimemez M, King NJ, Handelsman DJ, Allan CM. Sertoli and germ cell development in hypogonadal (hpg) mice expressing transgenic follicle-stimulating hormone alone or in combination with testosterone. Endocrinology. 2003 Feb; 144(2):509-17.
9. Setchell BP. Blood-testis barrier, junctional and transport proteins and spermatogenesis. AdvExp Med Biol. 2008;636:212-33
10. Colín A, Gomez-lópez N, Durna, Barroso G, Oehninger S. The effect of age on the expression of apoptosis biomarkers in human spermatozoa. FertilSteril 2011;94:2609-14.
11. Baker MA, Aitken RJ. Proteomic insights into spermatozoa: critiques, comments and concerns. Expert Rev Proteomics. 2009 Dec;6(6):691-705.
12. Avendaño C, Franchi A, Taylor S, Morshedi M, Bocca S, Oehninger S. Fragmentation of DNA in morphologically normal human spermatozoa. FertilSteril. 2009 Apr;91(4):1077-84.
13. Oehninger S. Molecular basis of human sperm-zonapellucida interaction. Cells Tissues Organs 2001;168:58-64.
14. Avendaño C, Franchi A, Oehninger S. Pregnancy-specific {beta}-1-glycoprotein 1 and human leukocyte antigen-E mRNA in human sperm: differential expression in fertile and infertile men and evidence of a possible functional role during early development.Hum Reprod. 2009 Feb;24(2):270-7.


